RNA
RNAの特徴
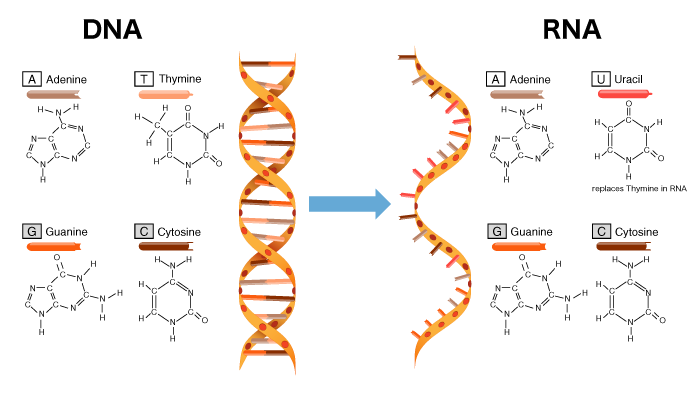
RNAはDNAと同じ核酸で、ヌクレオチドと呼ばれるリン酸・塩基・糖から成る基本構造を持ち、ヌクレオチドが連なった構造(ポリヌクレオチド)をとります。RNAは、転写により一部のDNA配列を鋳型として合成されます。DNAとRNAの違いは3つあります。まず、一本鎖のポリヌクレオチドであることです。二番目は、糖の種類です。RNAの糖(リボース)は、酸素分子がDNAの糖(デオキシリボース)より一つ多いことです。三番目は塩基の種類です。DNAの塩基はA、T、G、Cですが、RNAはTのかわりにUになります。
RNAの種類
RNAは、翻訳されるかどうかで大きく2種類に分けられます。翻訳を受けるRNAからは、タンパク質が合成されます。DNAからタンパク質をコードする配列をコピーして、タンパク質合成を担うリボソームまでDNAの遺伝情報を運ぶ役目を持つメッセンジャーRNA(mRNA)がこれに相当します。
一方、タンパク質をコードしない配列を持ち、翻訳を受けないRNAを総称してノンコーディングRNA(non-coding RNA; ncRNA)と呼びます。従来から知られているリボソームRNA(rRNA)やトランスファーRNA(tRNA)もncRNAに分類されます。RNA interference(RNAi)経路において機能的に重要な役割を担っているsmall ncRNA(miRNA、siRNA、piRNA)や、核内機能ドメイン構築への関与が多数報告されている長鎖のmRNA型ncRNAなどが注目を集めています。
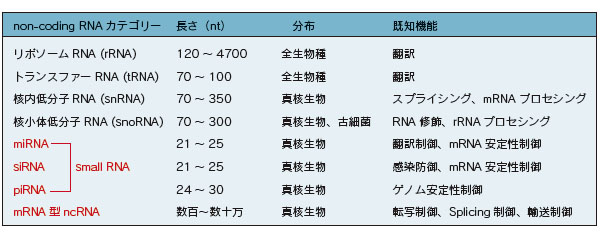
rRNA
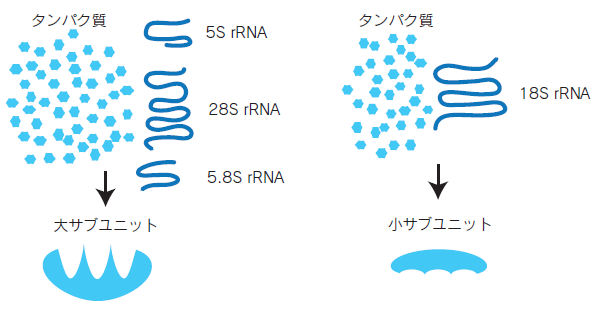
リボソームRNA (rRNA)は、翻訳装置として働くリボソームの構成因子です。リボソームは大小二つのサブユニットから成る巨大なRNA-タンパク質複合体です。rRNAは、触媒作用を持ち、翻訳時にアミノ酸が一つずつ結合していく際のペプチド結合の形成に関わることが知られています。
ゲノムにはrRNA遺伝子が多数存在します。rRNAはタンパク質を合成するリボソームに大量に含まれるので、ゲノム上にrRNAの領域が1つしかなかったら、最大速度で転写をしてもリボソームが不足するからです。細胞質には通常、数百万個のリボソームがあります。rRNAは一つながりの前駆体として転写され、真核生物では18S rRNA、5.8S rRNA、28S rRNA、5S rRNAという4種類に切断されます。その後200ヵ所以上の場所で修飾を受けますが、それにはsnoRNA(small nucleolar RNA)という小さなRNAが働いていると考えられています。このrRNAに多くのタンパク質が結合してリボソームという顆粒が形成されます。真核生物の80Sのリボソームは40Sの小サブユニット(18S rRNAと約30種類のタンパク質)と60Sの大サブユニット(5S rRNA、28S rRNA、5.8S rRNA と約50 種類のタンパク質)からできており、核の中の核小体という部分で作られます。核外に出て行った後、小胞体表面に結合して粗面小胞体を形成するものと、細胞質に残るものがあります。
tRNA
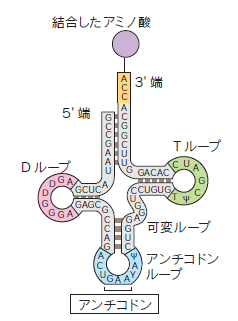 トランスファーRNA(tRNA)は、翻訳時にリボソームまでアミノ酸を運びます。クローバーリーフモデルと呼ばれる二次構造をとり、さらに折りたたまれてL型の構造をとっています。
トランスファーRNA(tRNA)は、翻訳時にリボソームまでアミノ酸を運びます。クローバーリーフモデルと呼ばれる二次構造をとり、さらに折りたたまれてL型の構造をとっています。
tRNAもまた翻訳時に大量に使用されるため、rRNAと同様にゲノム上にtRNA遺伝子が多数存在します。そこから一本の前駆体RNAが転写された後、切断され、修飾を受けて完成します。tRNAの修飾の種類は多く、現在までに100種類以上の修飾が同定されています。また、真核生物のtRNAの3'末端にはCCAという3つの塩基が付加されます。そこにアンチコドンに対応するアミノ酸が結合(チャージ)して機能します。
snRNA
核内低分子RNA(small nuclear RNA; snRNA)は、従来から知られているncRNAの一つです。U snRNAは、その配列中にウリジンを豊富に有し、タンパク質と複合体を形成してU snRNPとして機能します。U1、U2、U3、U4、U5、U6の6種類のU snRNPは、スプライソソーム構成因子としてスプライシングの調節に寄与します。この6種類のうち、U6 snRNAは、RNAポリメラーゼIIIの転写産物で5'末端にγ-メチルグアノシンキャップを有し、Lsmタンパク質群と結合しています。U6 snRNA以外はRNAポリメラーゼIIで転写されて5'末端に7-メチルグアノシンキャップ(m7G cap)を付加された後、核外に運ばれます。細胞質でSmタンパク質と結合し、m7G capがさらにメチル化されてトリメチルグアノシンキャップ(m3G cap)となり、再び核内へ運ばれた後でRNA修飾を受けることが知られています。
snoRNA
核小体低分子RNA(small nucleolar RNA; snoRNA)は、真核生物や古細菌で保存されていて核小体に存在するncRNAです。snoRNAにはU snRNAと同様にタンパク質と複合体を形成してsnoRNPとして機能します。snoRNAにはC/D snoRNAおよびH/ACA snoRNAの2つのグループに分けられます。これらの多くはrRNAの修飾やプロセシングに関与します。また、C/D snoRNAのうちU3 snoRNAはRNAポリメラーゼIIの転写産物で5'末端にm3G capを有することが知られています。
miRNA
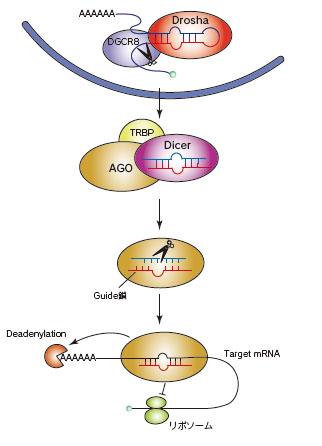 microRNA(miRNA)は、内在性の21-25塩基の短いRNAで、RNA interference(RNAi)経路を介して遺伝子発現制御に関与することが知られています。miRNAは、主にprimary-miRNA(pri-miRNA)としてゲノムから転写された後、核内でDroshaやDicerといったRNase III活性を有するタンパク質によるプロセシングを受け、21-25塩基の2本鎖RNAとなります。このようにしてできた2本鎖RNAはprecursor miRNA(pre-miRNA)と呼ばれ、細胞質へと輸送されます。
細胞質へ輸送された2本鎖miRNAは、RNA-induced silencing complex(RISC)に含まれるArgonaute(AGO)ファミリータンパク質に取り込まれます。その後片方のmiRNA(passenger 鎖、miRNA*と表記)が除去され、もう一方はmature miRNA(guide鎖)としてAGO内に残ります。AGOに取り込まれたmature miRNAの配列相補性を指標としてRISCが作用することにより、標的mRNAの翻訳抑制や分解といった遺伝子発現制御機構が働くと考えらえています。
microRNA(miRNA)は、内在性の21-25塩基の短いRNAで、RNA interference(RNAi)経路を介して遺伝子発現制御に関与することが知られています。miRNAは、主にprimary-miRNA(pri-miRNA)としてゲノムから転写された後、核内でDroshaやDicerといったRNase III活性を有するタンパク質によるプロセシングを受け、21-25塩基の2本鎖RNAとなります。このようにしてできた2本鎖RNAはprecursor miRNA(pre-miRNA)と呼ばれ、細胞質へと輸送されます。
細胞質へ輸送された2本鎖miRNAは、RNA-induced silencing complex(RISC)に含まれるArgonaute(AGO)ファミリータンパク質に取り込まれます。その後片方のmiRNA(passenger 鎖、miRNA*と表記)が除去され、もう一方はmature miRNA(guide鎖)としてAGO内に残ります。AGOに取り込まれたmature miRNAの配列相補性を指標としてRISCが作用することにより、標的mRNAの翻訳抑制や分解といった遺伝子発現制御機構が働くと考えらえています。
siRNA
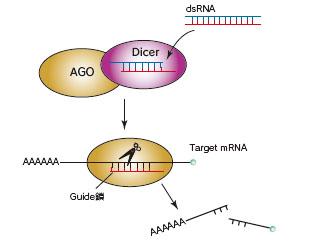 short interfering RNA(siRNA)は、ウイルス感染などの様々な要因で生じる長い2本鎖RNA(dsRNA)が由来とされています。Dicerにより、長いdsRNAが21-25 ntの2本鎖RNA(siRNAduplex)へと変換されます。siRNA duplexのうち、guide鎖がRISC(RNA-induced silencing complex)中のAGOに取り込まれ、silencerとして機能します。RISCに取り込まれなかったpassenger鎖は分解されます。
short interfering RNA(siRNA)は、ウイルス感染などの様々な要因で生じる長い2本鎖RNA(dsRNA)が由来とされています。Dicerにより、長いdsRNAが21-25 ntの2本鎖RNA(siRNAduplex)へと変換されます。siRNA duplexのうち、guide鎖がRISC(RNA-induced silencing complex)中のAGOに取り込まれ、silencerとして機能します。RISCに取り込まれなかったpassenger鎖は分解されます。
piRNA
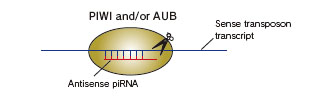 PIWI-interacting RNA(piRNA)は、トランスポゾンの転写産物の1本鎖RNAが由来とされ、DicerなどのRNase III活性を持つタンパク質によるプロセシングを必要としないprimary processing pathwayを介して生合成されます。生殖細胞特異的な発現が認められているPIWIタンパク質にローディングされることにより、トランスポゾンのsilencingに寄与していると考えられています。
PIWI-interacting RNA(piRNA)は、トランスポゾンの転写産物の1本鎖RNAが由来とされ、DicerなどのRNase III活性を持つタンパク質によるプロセシングを必要としないprimary processing pathwayを介して生合成されます。生殖細胞特異的な発現が認められているPIWIタンパク質にローディングされることにより、トランスポゾンのsilencingに寄与していると考えられています。
mRNA型ncRNAと核内機能ドメイン
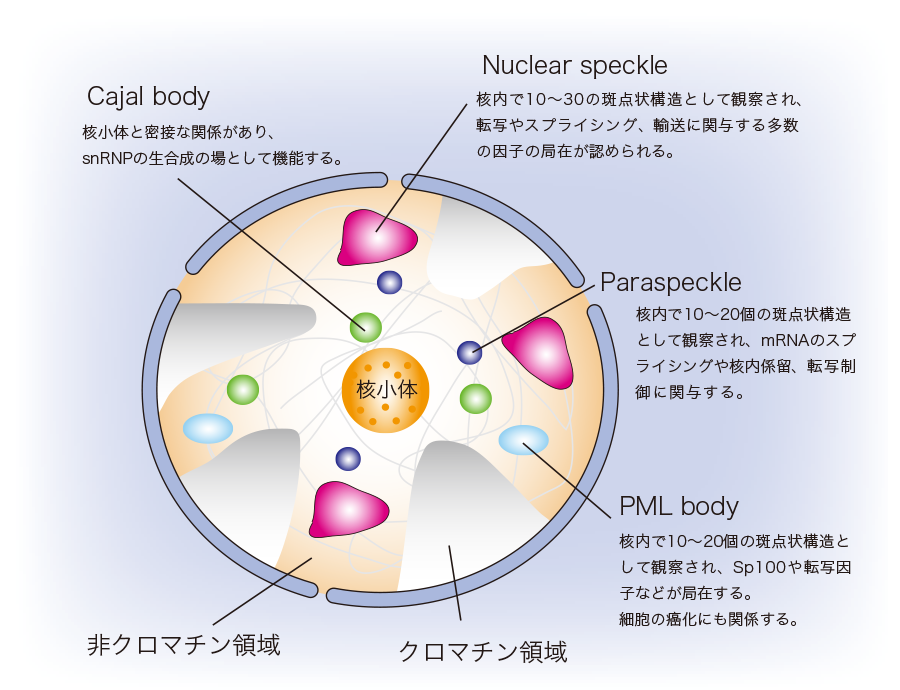
近年のトランスクリプトーム解析の発展により、ヒトにおいては、全ゲノム領域の9割以上が転写されているとの知見が得られています。また、ヒトの場合、RNAに転写されるゲノム情報の95%以上がncRNAであると見積もられています。つまり、転写されて生じるRNA の大部分はncRNAであるということになりますが、これらのncRNAの大半がアノテーションをつけられておらず、どのような機能を有しているのか理解されていないのが現状です。これらの中には、タンパク質はコードしないが転写後のプロセシングを受けるmRNA型ncRNAの存在も確認されています。 RNA研究が盛んになるにつれて、mRNA型ncRNAの大半は核内に留まり、様々な機能ドメイン構築に関与しているという事が次第に明らかとなってきました。ncRNAやRBPなどで構成される核内機能ドメインにおいては、機能単位ごとにncRNAやRBPが時空間的に制御・配列されてクラスター(集合体)を形成しており、MBLの提唱しているRiboClusterという概念を裏付けるものとして注目されています。
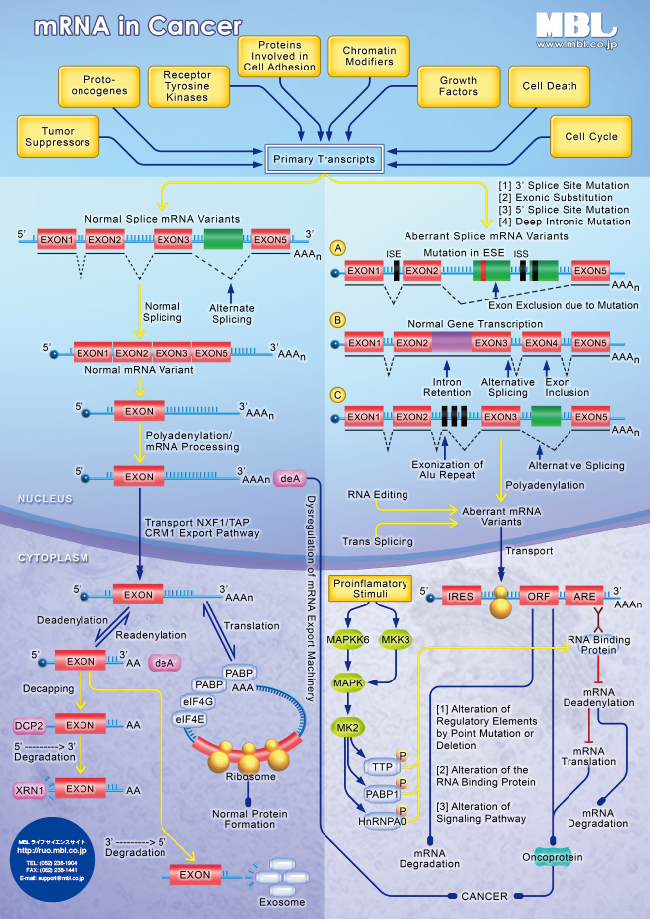
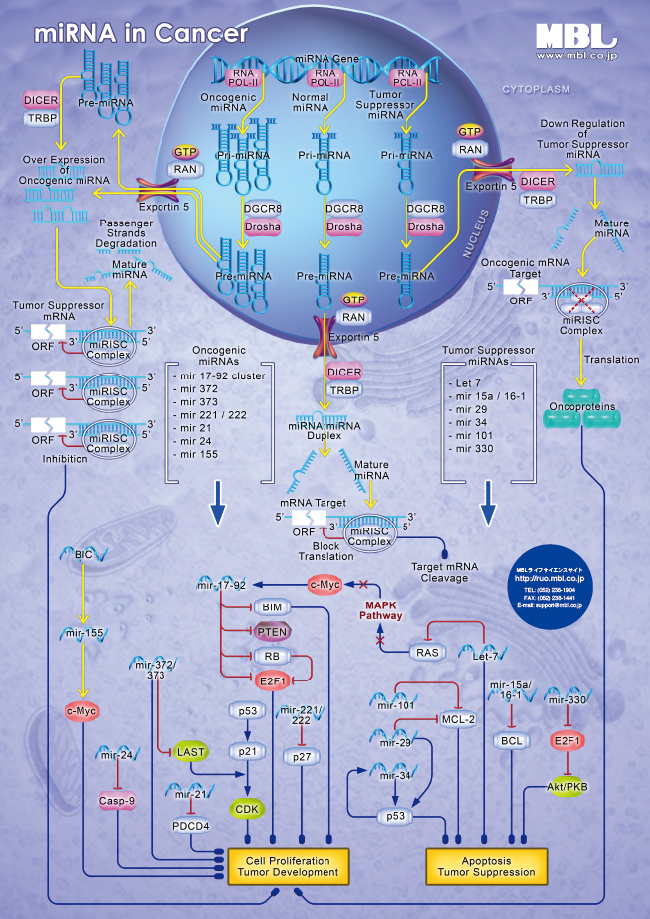
パスウェイポスター
| mRNA in Cancer PDF版(402KB) | miRNA in Cancer PDF版(392KB) |
 |
 |
 RIP-Assay Kit
RIP-Assay Kit